Gonorrhea Bacteria Feeds on Amino Acids
A Microbial Biorealm page on the genus Neisseria gonorrhoeae
Classification
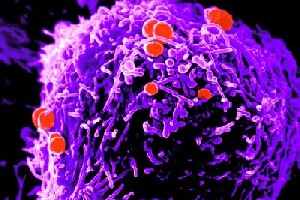
Neisseria gonorrhoeae. © I. Boulton and S. Gray-Owen Nat Immunol. 2002 3:229-236.
Higher order taxa
Domain: Bacteria
Phylum: Probacteria
Class: Beta Probacteria Order: Neisseriales
Family: Neisseriaceae
Species
Neisseria Gonorrhoeae
Description and significance
Neisseria gonorrhoeae is a gram-negative coccus, or bacteria whose overall shape is spherical. It is usually seen in pairs with adjacent sides flattened. The organism is usually found interacellulary in polymorphonuclear leukocytes, or a specific category of white blood cells with varying shapes of nuclei, of the gonorrhea pustular exudates [8] with humans as its only natural host. [10] N. gonorrhoeae is highly efficient in using transferrin-bound iron for in vitro growth. Many strains can also utilize lactoferrin-bound iron. The bacteria bind only human transferrin and lactoferrin. This specificity is thought to be the reason these bacteria are exclusively human pathogens. [8]
The bacterium was first discovered in 1879 by a German physician Albert Ludwig Sigesmund. [1] This organism is relatively fragile and is susceptible to temperature changes, drying, UV light, and some other environmental conditions. [8] The recommended procedure for isolating Neisseria gonorrhoeae involves the inoculation of a specimen directly onto a nutritive growth medium that is at room temperature and immediate incubation at 35-37ºC in an atmosphere of 3 – 10% added CO2. [6] Strains are inconsistent in their cultural requirements so the media needed for growth and isolation of the organism contain hemoglobin, NAD, yeast extract and other supplements.
Genome structure
Neisseria gonorrhoeae have a circular DNA genome. [7] N. gonorrhoeae strain 1090 genome was sequenced by the University of Oklahoma. [11] The genome length is 2,153,922 nt and contains 2069 genes and 67 structural RNAs. It also has 2002 protein genes. This includes the opacity (Opa) proteins which are responsible for the opaque colony phenotype caused by tight junctions between adjacent Neisseria, and are also responsible for tight adherence to host cells. This organism is also naturally competent for the update of DNA. [1] Neisseria gonorrhoeae can produce one or several Opa proteins. These proteins are subject to phase variation and are usually found on cells from colonies possessing a unique opaque phenotype called O+. At any particular time, the bacterium can express zero, one, or several different Opa proteins, and each strain has 10 or more genes for different Opas. [8] More specifically, during infection, N. gonorrhoeae is like to encounter hydrogen peroxide, which inhibits growth. Since it is an obligate human pathogen, it would not be exposed to typical environmental stress such as UV light, ionizing radiation, or chemical mutagens. The type of DNA damage N. gonorrhoeae would come across is oxidative. [12]
N. gonorrhoeae genome contains many genes that are predicted to be involved severeal DNA repair pathyways. Recombinational DNA repair has been studied in N. gonorrhoeae and requires the recA and recX genes, which act with either the RecBCD pathway (recB, recC, and recD genes) or the RecF-like pathway (recO, recQ, recR, and recJ genes). Also, contributing to the recombinational DNA repair pathway is the Holliday junction processing enzymes encoded by recG, ruvA, ruvB, and ruvC. N. gonorrhoeae seems to use both DNA recombinational repair pathways simultaneously. This is in contrast to Escherichia coli, where mutants in the RecF pathway generally show phenotypes only in the context of recBC sbcBC mutations. This leads to the conclusion that recombinational DNA repair is especially important for the repair of damaged DNA in N. gonorrhoeae. [12]
During repair of oxidatively damage in E.coli, recA and other reocombinational rpair genes have been shown to be important. E. coli recA is important for both functions in DNA repair and its role in the induction of the SOS response of DNA repair. But because N. gonorrhoeae does not have SOS response, it does not use recA for the repair of oxidatively damaged DNA. [12]
Analysis has shown that only recN, a single known DNA repair and recombination gene is upregulated after hydrogen peroxide treatment. It is unclear as to what the exact role of this gene is, but it seems to function in the repair of DNA double strand breaks. In addition, an N. gonorrhoeae recN mutant displays decreased survival to nalidixic acid and hydrogen peroxide, both of which can result in DNA double-strand breaks. [12]
Although several gonococcal genes have been identified that protect against oxidative damage, few of them are predicted to function in the repair of DNA. To date, only two genes that are involved in DNA repair and recombination have been found to protect against oxidative damage in N. gonorrhoeae. Both the N. gonorrhoeae recN mutant and a mutant inactivated in priA, which is involved in replication restart, show decreased resistance to oxidative damaging agents. In contrast to E. coli recA, N. gonorrhoeae recA was reported to not protect against oxidative damage caused by H2O2. This suggests that DNA repair and recombination enzymes may differ between N. gonorrhoeae and E. coli in their importance to the repair of oxidatively damaged DNA. [12]
RecA, genes of the RecBCD and RecF-like recombination pathways, and genes whose products are involved in Holliday junction processing are all important for mediating repair of oxidative damage. Futhermore, data suggest that these genes are expressed at basal levels sufficient to mediate repair and do not need to be upregulated upon encountering DNA damage in order to function in N. gonorrhoeae.
The recent demonstration that N. gonorrhoeae is polyploid suggests that, in the event of chromosomal damage, these additional copies of the chromosome could provide the genetic information present on the damaged copy, perhaps anticipating the necessity of recombinational repair. Therefore, of the many mechanisms of resistance used by N. gonorrhoeae to combat oxidative insult, recombinational DNA repair appears to be one layer of resistance. [12]
Cell structure and metabolism
Neisseria gonorrhoeae posses a typical gram negative outer membrane that is composed of proteins, phospholipids, and lipopolysaccharide (LPS). Neisserial LPS is unique in that it has highly-branched basal oligosaccharide structure and the absence of repeating O-antigen subunits. Thus, they are referred to as lipooligosaccharide (LOS). During growth, the bacterium releases outer membrane fragments called "blebs". These contain LOS and may have a role in the pathogenesis if they are distributed during the course of an infection. [8]
The bacterium have fimbriae, which is a proteinaceous appendage that is thinner than a flagellum. They play a major role in adherence and extend several micrometers from its cell surface. [8] There are four types of N. gonorrhoeae based on the presences of fimbriae and they are called T1, T2, T3, and T4. [4]. In vitro studies show that these piliated cells bind more efficiently to eukaryotic cells than non piliated cells, which suggests that the pilus structure plays an important role in this interaction [3]
N. gonorrhoeae also can move in a jerky fashion across solid surfaces. This type of motility is called twitching, which depends on type IV pili and takes place by a "grappling hook" mechanism, which is the extension of the pilus, its attachment, and its retraction back into the cell. Twitching motility also contributes to the formation of biofilms. [9] During growth, Neisseria gonorrhoeae releases soluble fragments of peptidoglycan. These molecules are implicated in the pathogenesis of different forms of gonococcal infection. A major peptidoglycan fragment released by N. gonorrhoeae is identical to the tracheal cytotoxin of Bordetella pertussis and has been shown to kill ciliated fallopian tube cells in organ culture. In the examination of the role of other putative lytic transglycosylases in peptidoglycan-derived cytotoxin (PGCT) production, results suggest that this gonococcal gene (ltgA) encodes a lytic peptidoglycan transglycosylase and that it is responsible for a significant proportion of the PGCT released by N. gonorrhoeae .[2]
N. gonorrhoeae genome contains homologues of enzymes involved in PG recycling, and the levels of turnover are consistent with a certain level of recycling occurring in gonococci. [2] It is unknown if N. gonorrhoeae have cytoplasmic proteins for sensing PG fragments; but, this would be a favorable mechanism for controlling cell processes, including autolysis.
The presence of two, maybe more enzymes with potentially redundant functions either indicates that gonococci have an elaborate backup system for cell wall processes or may suggest that the enzymes have different functions or are differently regulated or localized. AtlA is encoded in a group of type IV secretion genes in the gonococcal genetic island, and recent evidence suggests that AtlA may have a role in assembly of the type IV secretion system. [2]
PGCT is expected to be released during infection, due to the extensive turnover and release of PG fragments in vitro. [2] Although many lytic transglycosylases were characterized in E. coli, the genes for PGCT production have not been previously characterized in bacteria in which PGCT is thought to act in infection.
These organisms are aerobic, strongly oxidase-positive, have an oxidative metabolism, are susceptible to drying and are fastidious (growth is inhibited by free fatty acids). [4]
Ecology
Neisseria gonorrhoeae is the cause of the sexually transmitted disease gonorrhea. It natural environment is in the human body and it only known for its pathogenic characteristic. Neisseria gonorrhoeae is not used for any known biotechnology.
Pathology
Patheogenesis
Neisseria gonorrhoeae infections are acquired by sexual contact and usually affect the mucous membranes of the urethra in males and the endocervix and urethra in females. The pathogenic mechanism involves the attachment of the bacterium to nonciliated epithelial cells through pili (fimbriae) and the production of lipopolysaccharide endotoxin. [8]. Neisseria gonorrhoeae is only found after sexual contact with an infected person (or in the case of infections in the newbord, direct contact).
Adherence is mainly done through fimbriae and opa (P.II) protein although nonspecific factors such as surface charge and hydrophobicity may play a role. The bacteria only attach to microvilli of nonciliated columnar epithelial cells and not ciliated cells. [8]
After the bacteria attach to the nonciliated epithelial cells of the fallopian tube, they are surrounded by the microvilli that draw them to the surface of the mucosal cell. Then the bacteria enter the epithelial cells by a process called parasite-directed endocytosis. During this process, the membrane of the mucosal cell retracts and pinches off a membrane-bound vacuole that contains the bacteria. The vacuole is transported to the base of the cell, where the bacteria are released by exocytosis into the subepithelial tissue.
During infection, bacterial lipooligosaccharide (LOS) and peptidoglycan are released by autolysis of cells. Both bacterial polysaccharides activate the host's alternative complement pathway, while LOS stimulates the production of tumor necrosis factor (TNF) which causes cell damage. Neutrophils are then attracted to the site and feed on the bacteria. For reasons not known, many gonococci are able to survive inside of the phagocytes. [8] Gonococcal LOS produces mucosal damage in fallopian tube organ cultures and brings about the release of enzymes, such as proteases and phospholipases Thus, gonococcal LOS seems to have an indirect role in mediating tissue damage.
Sometimes Neisseria gonorrhoeae can enter the bloodstream causing a Gram-negative bacteremia which may lead to a disseminated bacterial infection. Strains of N. gonorrhoeae that cause disseminated infections are usually resistant to complement and the serum bactericidal reaction. This accounts for their ability to persist in the bacteria infected blood. Gram-negative bacteremias of this kind can be aggravated by the lysing of bacterial cells which may simply liberate soluble LPS. [8]
Virulence Factors
Although it does not produce any exotoxins, Neisseria gonorrhoeae has a wide range of virulence determinants. The first stages of infection, which includes adherence and invasion, are mediated by surface components. The bacterium first attaches to epithelial cells by means of its fimbriae, specifically N-methylphenylalanine (Type IV) pili, with the main subunit PilE. After initial attachment, the bacteria enter a second stage of binding mediated by the outer membrane protein P.II (also known as Opa) which is needed for tight binding and invasion of epithelial cells. Also, P.II from one bacterium will bind to LOS of an adjacent bacterium, which allows for the construction of a small colony that may function similarly to a biofilm. Neisseria gonorrhoeae also produces an IgA1 protease that may take part in the colonization stage. [8]
The outer membrane porin of N. gonorrhoeae P.I (also known as Por) is equivalent to the ompC and ompF porins of E. coli. They are involved in the passage of solutes through the outer membrane. However, P.I apparently has a role in virulence that allows the gonococci to survive inside of phagocytes. Purified P.I has also been shown to inhibit the ability of phagocytes to kill ingested bacteria. [8] The lipooligosaccharide (LOS) is thought to be responsible for most of the symptoms of gonorrhea. Gonococcal LOS triggers an intense inflammatory response. The activation of complement, attraction and feeding by phagocytes, and the lysing of the phagocytes themselves, contributes to the purulent discharge. The local production of TNF is thought to be the main cause of damage to the fallopian tubes. In addition, in strains that cause systemic infection, LOS binds sialic acid from the serum forming a microcapsule of sialylated LOS, which allows the gonococci to resist the host immune response and serum bactericidal reaction. [8]
Nonsialyated LOS and P.I (Por) on the bacterial surface are known to be effective targets for bactericidal antibodies. However, if antibodies produced against P.III (also known as Rmp) react with their antigenic site on the gonococcal surface, the effect is to block bactericidal antibodies against LOS and P.I and to protect the bacterium from complement-mediated lysis.
Neisseria gonorrhoeae also have a well-developed iron acquisition system that allows it to extract iron from its host during growth, which is necessary to support bacterial invasion. The bacterium is able to form two transferrin receptors (Tbp1 and Tbp2) and one lactoferrin receptor (Lbp) in its outer membrane, which are stimulated under low-iron conditions, and are able to directly extract iron from transferrin and lactoferrin. These proteins can also extract iron from heme and hemoglobin. [8]
Neeisseria gonorrhoeae usually infects the mucous membranes causing infections such as urethritis, cervicitis, salpingitis, pelvic inflammatory disease, proctitis, conjunctivitis and pharyngitis. [10]
Symptoms
In males there is an approximate 2-3 day incubation period after which a purulent discharge from the urethra and dysuria develops. Around 95% of infected males are symptomatic. Rare complications include prostatitis, epididymitis, and periurethral abcesses. [10]
In women, Neisseria gonorrhoeae primarily infects the cervix in women. The symptoms of gonorrhea are often mild and most women who are infected do not have symptoms. Even when a woman has symptoms, they can be so non-specific and can be mistaken for a bladder or vaginal infection. Symptoms include vaginal discharge, dysuria, and abdominal pain. Around 10%-20% of infected women develop these complications. In 1%-3% of infected women and a lower percentage of infected men the bacterium disseminates via the blood causing bacteremia and arthritis. [10]
Application to Biotechnology
Neisseria gonorrhoeae is the cause of the sexually transmitted disease gonorrhea. It natural habitat is in the human body and it only known for its pathogenic characteristic. Neisseria gonorrhoeae is not used for any known biotechnology.
Current Research
N terminus determinants of MinC from Neisseria gonorrhoeae mediate interaction with FtsZ but do not affect interaction with MinD or homodimerization
In the present study, interaction between the cell division inhibitor MinC from Niesseria gonorrhoeae (MinCNg) and the gonococcal cell division proteins MinDNg and FtsZNg are demonstrated. Protein truncation and site-directed mutagenic approaches will determine which N-terminal residues were essential for cell division inhibition by MinCNg. Truncation from or mutation at the 13th amino acid of the N terminus of MinCNg resulted in loss of protein function. The bacterial two-hybrid system showed that the interaction of MinC Ng with FtsZ Ng is annulled with the truncation of 13 N-terminal residues. It demonstrated that MinC Ng truncation at the 13th leucine residue lost its ability to interat with FtsZ Ng, displaying a reason fro loss of functionality upon N terminal truncation.
The data confirms interaction among gonococcal cell division proteins and determind the necessity of the 13th amino acid for MinC Ng function. This study demonstrates that the N-terminus of MinC from N. gonorrhoeae is involved in protein functionality and interaction with FtsZ Ng. Delection of 13 amino acids from the N terminus or point mutation of the 13th residue of MinC Ng renders it non-functional as an inhibitor of cell division. This method demonstrated the 13th amino acid of MinC Ng - FtsZ Ng interaction is essential for gonococcal cell division. [5]
Weak Linkage in parC of Neisseria gonorrhea: Identification of Mutation Prone Point that can Lead to Drug Resistance
Mutation of N. gonorrhoeae pathogen is believed to be an important factor that can initiate drug resistance. In fluoroquinoline-resistant N. gonorrhoeae, the amino acid mutations in the fluoroquinoline-resistant detemining region (QRDR) of the parC gene are an important factor. Here, the author performed a bioinofrmatic analysis to study the positions that tend to comply peptide motifs in the amino acid sequence of parC. To identify the weak linkage in parC, a new bioinformatics tool called GlobPlot was used.
According to this work, positions 1–11, 80–86, 107–120, 147–160, 217–226, 288–294, 584–597 are identified as the position resistance to mutation. Based on this study, the weak linkages in the parC can be identified and generate information for the expectation of possible new mutations that can lead to failure of gonorrhea therapy. In addition, the results from this study provide information for further research on the diagnosis for mutant gonorrhea and new drug development. [14]
Multiple doses of cefodizime are necessary for the treatment of Neisseria gonorrhoeae pharyngeal infection
A single dose of cefodizime (CDZM), ceftriaxone(CTRX), or spectinomycin (SPCM) is what is recommended for the treatment of gonococcal urethritis or uterine cervicitis in the era of multidrug-resistant Neisseria gonorrhoeae; namely, cefozopran-resistant N. gonorrhoeae (CZRNG). It was previously found that a single dose of CDZM completely eradicated multidrug-resistant N. gonorrhoeae in patients with urethritis and uterine cervicitis, and so a single 1.0-g dose of CDZM for the treatment of N. gonorrhoeae pharyngeal infection, including infections with CZRNG was tried.
The eradication rate of N. gonorrhoeae from the pharynx was 63.0% with a single 1.0-g dose of CDZM, while the rate for CZRNG with the same dose of CDZM was 38.5%. N. gonorrhoeae was completely eradicated from the pharynx when patients received one or two additional doses of CDZM. It was found that N. gonorrhoeae was completely eradicated from the pharynx when patients received one or two additional doses of CDZM. Therefore, it can be concluded that two to three doses of CDZM are necessary for the treatment of CZRNG pharyngeal infection. [13]
References
1. "Albert Ludwig Sigemund Neisser: discoverer of the cause of gonorrhea", s Baylor College of Medicine, Department of Pediatrics, Houston, Texas
2. Cloud, Karen A., Dillard, Joseph P., A Lytic Transglycosylase of Neisseria gonorrhoeae Is Involved in Peptidoglycan-Derived Cytotoxin Production, Department of Medical Microbiology and Immunology, University of Wisconsin Medical School, American Society for Microbiology, 2002
3. Deal, C.D., Sromberg, N., Nyberg, G., Normark, S., Karlsson K.A., So, M. Department of Medical Biology, Research Institiute of Scripps Clinic, Pilin independent binding of Neisseria gonorrhoeae to immobilized glycolipids, Martinus Nijhoff Publishers 1987.
4. Fix, Douglas F., Neisseria, Department of Microbiology, Southern Illinois University at Carbondale. 2007.
5. Greco-Stewart, V., Ramirez-Arcos, S., Liao, M., Dillon, J.R., N terminus of MinC from Neisseria gonorrohoeae mediate interaction with FtsZ but do not affect interaction with MinD or homodimerization. Springer-Verlag 2007
6. Kaye D. Evans, Ellena M. Peterson, Janis I. Curry, J.R. Greenwood, Luis M. De La Maza. "Effect of Holding Temperature on Isolation of Neisseria gonorrhoeae" Journal of Clinical Microbiology, Dec 1986, American Society for Microbiology, p. 1109-1110
7. KEGG: Kyoto Encyclopedia of Genes and Genomes, Neisseria gonorrhoeae. Kanehisa Laboratories 1995-2007
8. Kenneth Todar, University of Wisconsin-Madison. Department of Bacteriology. 2004
9. Moselio Schaechter, John L. Ingraham, Frederick C. Neidhardt, Microbe ASM Press, American Society for Microbiology, Washington DC, 2006 pp.268
10. NCBI: Neisseria Gonorrhoeae, Accessed August 23, 2007
11. "Neisseria gonorrhoeae causes Gonorrhea" European Bioinformatics Institue 2006-2007, European Molecular Biology Laboratory.
12. Neisseria gonorrhoeae, Los Alamos National Laboratory, University of California for the UC Department of Energy, 1997.
13. Stohl, Elizabeth A, Seifert H. Steven. Neisseria gonorrhoeae DNA Recombination and Repair Enzymes Protect against Oxidative Damage Caused by Hydrogen Peroxide, Northwestern University, Feinberg School of Medicine, Illinois, American Society for Microbiology. 2006
14. Tetsuro Matumoto, Testuro Muratani, Kocihi Takahashi, Toshihiro Ikuyama, Diasuke Yokoo, Yukiko Ando, Yumiko Sato, Motoko Kurashima, Hiroshi Shimokawa, Shigeaki Yanai, Multiple doses of cefodizime are necessary for the treatment of Neisseria gonorrhoeae pharyngeal infection., Japanese Society of Chemotherapy and The Japanese for Infectious Diseases, 2006
15. Wiwanitit, Viroj, Weak Linkage in parC of Neisseria Gonorrhoeae: Identification of Muetation Prone Point that can Leave to Drug Resistance, Springer Sciences + Business Media, 2007.
Edited by Maricor Sicat, student of Rachel Larsen
Edited by KLB
Source: https://microbewiki.kenyon.edu/index.php/Neisseria_gonorrhoeae
0 Response to "Gonorrhea Bacteria Feeds on Amino Acids"
Enregistrer un commentaire